Someone plants a ginkgo seedling from a nursery. The tree grows beautifully for twenty years: clean pyramidal form, fan-shaped leaves that turn gold every autumn, never a pest problem. Then one October the tree drops a crop of plum-sized fruit that smells, according to one botanist, like raw dog vomit. The neighbors complain. The arborist who gets the call has to explain that this is a female tree and there is nothing to be done about it now except remove and replant. Twenty years, because nobody understood how that tree reproduces.
Or consider the homeowner who plants a single Fuji apple tree and waits five years for a harvest that never comes. The tree blooms beautifully every spring. Bees visit. But no fruit sets, because Fuji cannot pollinate itself and there is no compatible partner within bee range.
Both problems trace back to the anatomy of flowers. Not the showy petals you photograph in spring, but the reproductive machinery hidden inside them: which parts make pollen, which parts receive it, and whether a given tree carries both or only one. This is the guide that makes those distinctions practical.
The previous guides in this series covered the macroscopic structure of trunks, branches, and twigs, the wood and bark vascular system, leaf anatomy, root architecture, and mycorrhizal partnerships. All of those structures exist, ultimately, to support reproduction. This guide covers the reproductive structures themselves.
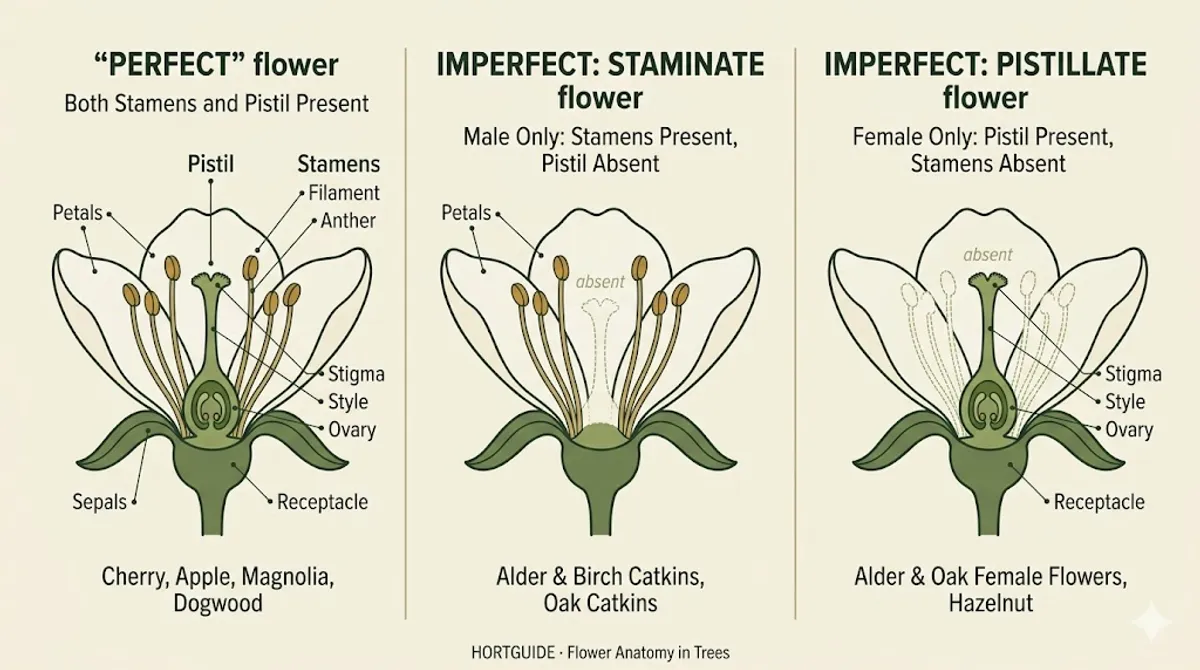
The Four Parts of a Flower
Every angiosperm flower is built from the same four sets of parts, arranged in concentric rings called whorls. Start from the outside and work in.
Sepals form the outermost whorl, collectively called the calyx. These are the green, leaf-like parts that enclose and protect the flower bud before it opens. On a magnolia bud in early spring, the tough outer scales you peel away are sepals. On a rose, the green star at the base of the flower is the calyx. Sepals are structural: they protect the developing flower parts inside, then fold back out of the way when the flower opens.
Petals form the next whorl inward, collectively called the corolla. These are the parts you notice. Petals are the showy, often colorful structures that attract pollinators with color, pattern, and sometimes ultraviolet markings invisible to humans but visible to bees. Not every flower has showy petals. Oak flowers, alder catkins, and most conifer structures lack them entirely, for reasons that matter (see the pollination section below).
Stamens form the third whorl, collectively called the androecium. These are the male reproductive organs. Each stamen has two parts: a thin stalk called the filament and a pollen-producing head called the anther. When the anther matures, it splits open and releases pollen grains. If you have ever brushed against a lily and gotten yellow powder on your shirt, you touched an anther. On trees, the number of stamens per flower varies enormously: cherry blossoms have 20 to 30, magnolias may have over 100.
The pistil (or pistils) occupies the center of the flower, forming the gynoecium. This is the female reproductive organ. It has three parts: the stigma (a sticky or feathery tip that catches pollen), the style (a tube connecting the stigma to the ovary), and the ovary (the swollen base containing one or more ovules). After pollination, the ovary develops into the fruit. Every cherry you eat, every apple you pick, every maple samara that helicopters onto your deck was once an ovary inside a flower.
A flower with all four parts present is called complete. Magnolia, cherry, apple, and dogwood all have complete flowers. A flower missing any one of those four whorls is incomplete. This distinction sounds academic until you realize that most wind-pollinated tree flowers are incomplete: they lack showy petals because they have no pollinators to attract.
Perfect, Imperfect, and Why It Matters
Completeness describes whether all four whorls are present. A separate classification describes which sex organs the flower carries, and this one has direct consequences for your yard.
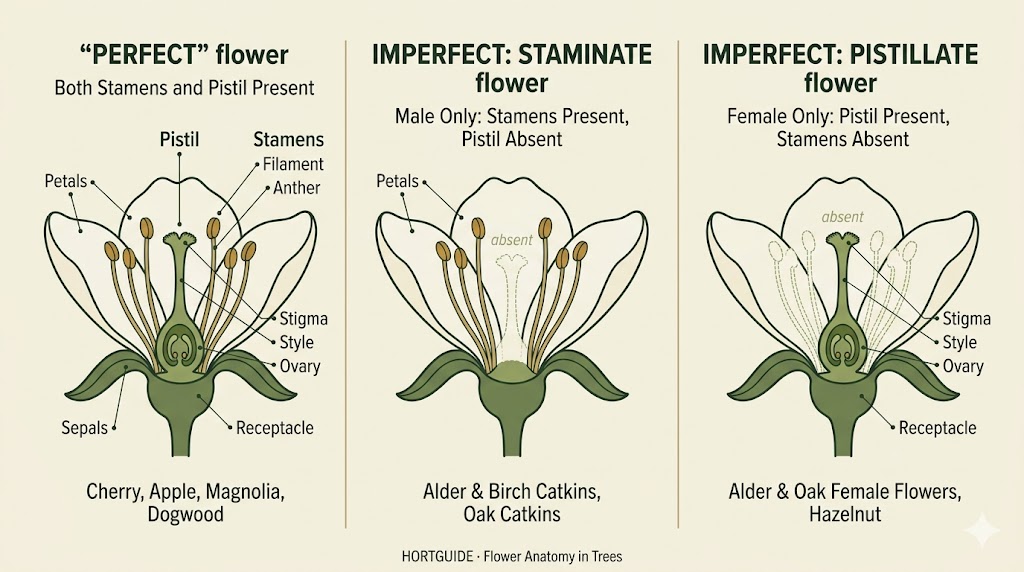
A perfect flower contains both stamens (male) and at least one pistil (female) in the same flower. Cherry, apple, magnolia, and most fruit trees have perfect flowers. Each individual blossom carries both sexes. This does not necessarily mean the tree can pollinate itself (self-incompatibility is a separate issue), but it does mean every flower has the potential to produce fruit.
An imperfect flower carries only one sex. A flower with stamens but no pistil is staminate (male). A flower with a pistil but no stamens is pistillate (female). Red alder catkins, the dangling structures that dust your car with yellow powder every February in the Puget Sound lowlands, are staminate flowers. The small, woody, cone-like structures on the same alder branch are the pistillate flowers that will become the alder “cones” you see in autumn.
Here is where the practical consequences begin. A tree species with imperfect flowers must answer a logistical question: where do the male and female flowers live?
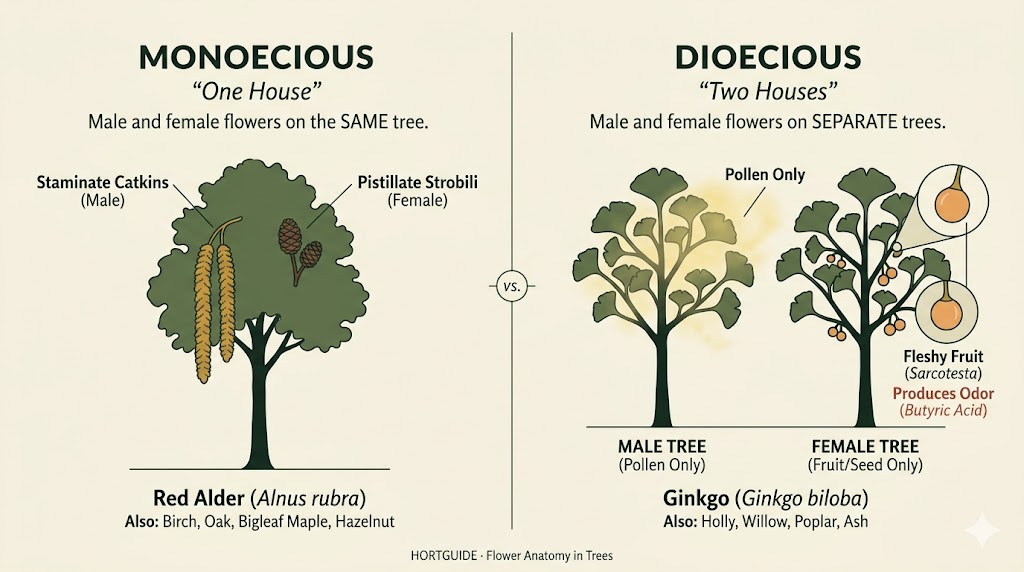
Monoecious trees carry both male and female flowers on the same individual plant. Red alder, birch, oak, and bigleaf maple are monoecious. A single tree has everything it needs to produce seed (though cross-pollination with another tree of the same species often improves seed set). Monoecious trees typically bear their male flowers in catkins or clusters positioned above or separate from the female flowers, which reduces self-pollination and encourages cross-pollination by wind or insects.
Dioecious trees carry male and female flowers on entirely separate individuals. A male ginkgo produces only pollen. A female ginkgo produces only seeds. They are different trees, the way a rooster is not a hen. Holly, willow, poplar, ash, and persimmon are also dioecious. This has three consequences you will encounter in practice:
-
A female dioecious tree cannot produce fruit or seed unless a male tree of the same species is close enough for pollen transfer. If you want holly berries for winter interest, you need at least one male holly within pollinator range of your female plants.
-
A male dioecious tree will never produce fruit. This is why nurseries sell named male cultivars of ginkgo (Autumn Gold, Princeton Sentry, Magyar) and male cultivars of ash. You are buying a guarantee that the tree will not produce the messy or malodorous fruit that female trees do.
-
If you plant an unselected seedling of a dioecious species, you may not know its sex for a decade or more. Ginkgo seedlings may take 20 years to flower. By then, removing and replacing a mature female tree is a significant expense.
A few trees complicate the clean monoecious/dioecious split. Red maple (Acer rubrum) is technically polygamo-dioecious: individual trees tend to be mostly male or mostly female, but some flowers on any given tree may be perfect. This is one reason “guaranteed male” red maple cultivars occasionally produce some fruit. Bigleaf maple (Acer macrophyllum) is similarly fluid: usually simplified as monoecious in field guides, but classified as polygamous in the USDA Plants Database, with staminate, pistillate, and occasionally perfect flowers borne in the same raceme. Biology does not always respect nursery catalog claims.
How Gymnosperm Cones Differ
Everything above applies to angiosperms, the flowering plants. But roughly a third of the trees in a typical Puget Sound lowland landscape are gymnosperms: Douglas-fir, western red cedar, pines, spruces, true firs, and hemlocks. These trees do not produce flowers at all. They reproduce using cones.
The distinction is fundamental. “Angiosperm” means enclosed seed: the ovules are protected inside an ovary, which develops into fruit. “Gymnosperm” means naked seed: the ovules sit exposed on the surface of a cone scale, with no ovary enclosing them.
Gymnosperms produce two types of cones. Male pollen cones (botanically, microsporangiate strobili) are small, soft, and often clustered near branch tips. They release clouds of pollen in spring and then disintegrate. If you have ever seen a yellow dusting on your car in April and traced it to your Douglas-fir or pine, that was pollen from male cones. Female seed cones (megasporangiate strobili) are the woody structures most people think of when they hear “pinecone.” These are larger, persist on the tree much longer, and contain the ovules that, after pollination, develop into seeds.
Pollination in gymnosperms is exclusively wind-driven. There are no petals, no nectar, no insect pollinators. Female cones secrete a sticky liquid called a pollination drop through a tiny opening in each ovule (the micropyle) to capture airborne pollen grains. The timeline from pollination to mature seed is much longer than in most angiosperms: in pines, it can take two to three years.

Douglas-fir illustrates the cone distinction clearly. The female cones are the familiar woody structures with three-pointed bracts protruding between the scales, the feature that makes Douglas-fir cones instantly identifiable. The male cones are small, reddish, and clustered among the needles in spring, rarely noticed by anyone who is not looking for them. Western red cedar’s male cones are even smaller, appearing as tiny reddish-brown tips on branch ends.
Ginkgo occupies a strange middle ground. It is a gymnosperm (its seeds are “naked,” not enclosed in an ovary), but its reproductive structures do not look like cones. Male ginkgos produce catkin-like pollen structures about an inch long, with loosely arranged stamens. Female ginkgos produce long-stalked structures with two opposing ovules. The fleshy, plum-shaped covering that develops around the seed is not a fruit (there is no ovary), but a structure called the sarcotesta. It is this sarcotesta that produces the notorious odor. Beneath it sits the hard seed shell (sclerotesta), and inside that, the edible “nut” prized in East Asian cuisine. Ginkgo is the only surviving member of a plant lineage that dates back roughly 150 million years, and its reproductive biology reflects that ancient origin.
Pollination: Wind, Insects, and What It Means for Your Yard
Trees use two primary strategies to get pollen from one flower to another, and the strategy a tree uses explains a surprising amount about its impact on your landscape.
Wind-pollinated trees invest in quantity. Because wind is random, the only way to ensure some pollen reaches a receptive flower is to produce enormous amounts of it. A single birch catkin can release millions of pollen grains. Red alder, the most common deciduous tree in Western Washington lowlands, flowers in February and March, well before its leaves emerge, so the canopy is open and pollen can travel freely. Wind-pollinated flowers are characteristically inconspicuous: no showy petals (nothing to attract), no nectar (no insect to reward), and exposed anthers or stigmas designed to catch the breeze. Catkins, the dangling clusters you see on alders, birches, oaks, and hazelnuts, are the signature form of wind-pollinated flowers.

This strategy is why you have allergies. The pollen counts that spike February through May in the Puget Sound region are almost entirely from wind-pollinated trees: red alder, birch, western red cedar, bigleaf maple, and oak. These trees flood the air with lightweight pollen designed to travel on currents. Insect-pollinated trees produce far less pollen, and their grains are heavier and stickier, built to adhere to a bee, not to float through a window screen.
Insect-pollinated trees invest in advertising. Showy petals, bright colors, fragrance, and nectar are all strategies to attract pollinators: bees, butterflies, beetles, flies. Each pollinator visit transfers a small, targeted payload of pollen, so the tree needs less total pollen but more investment per flower. Cherry blossoms, apple blossoms, magnolia flowers, and linden flowers are all designed for insect visitors. In the Puget Sound lowlands, native mason bees (Osmia lignaria) emerge in late March, timed precisely with early fruit tree bloom, and are among the most effective pollinators of cherries, plums, and apples in home orchards.
Self-incompatibility adds a complication for gardeners. Many fruit trees with perfect flowers (both sexes in every blossom) still cannot pollinate themselves. Apple, pear, sweet cherry, and most Japanese plums are self-incompatible: the pollen from one tree’s flowers is chemically rejected by the pistils of flowers on the same tree. The tree recognizes its own pollen and blocks fertilization. This is an evolutionary strategy to promote genetic diversity, but for a homeowner with a single apple tree, it means zero fruit. You need a second, compatible cultivar within bee range (roughly 100 feet, though some guidelines suggest up to 300 feet for adequate cross-pollination). Compatibility is specific: not any two apple varieties will cross-pollinate each other. Pollination charts exist for apples, pears, and cherries because the matchmaking matters.
From Flower to Fruit
After a pollen grain lands on a receptive stigma and grows a pollen tube down through the style to reach the ovule, fertilization occurs and the ovary begins developing into fruit. The type of fruit a tree produces follows directly from the flower anatomy that made it.
Drupes have a fleshy outer layer surrounding a single hard pit (the endocarp) that encloses the seed. Cherry, plum, peach, and the fruit of the native Cascara (Frangula purshiana) are all drupes. The pit is the hardened inner ovary wall.
Pomes are the characteristic fruit of the rose family: the fleshy part you eat develops from the receptacle (the base that held the flower parts), not from the ovary wall directly. Apple and pear are pomes. Cut an apple crosswise and the star-shaped seed cavity at the center is the actual ovary; everything surrounding it grew from accessory tissue.
Samaras are winged, dry fruits designed for wind dispersal. Bigleaf maple (Acer macrophyllum) produces the largest samaras of any native maple, paired wings that helicopter down from the canopy starting in September. Each samara contains one seed. The wing is a modified ovary wall. Ash trees also produce samaras, though single-winged rather than paired.
Acorns are the nuts of oaks, each one a single seed sitting in a scaly cup (the involucre, formed from modified bracts, not from the ovary). Oaks are wind-pollinated and monoecious. The pollen catkins appear in spring; the acorns mature by autumn, sometimes taking two years in red oak species.
Capsules like the long pods of northern catalpa (Catalpa speciosa) split open at maturity to release seeds. The dry, rattling pods that persist on catalpa through winter are dehiscent capsules.
Aggregate fruits like the cone-shaped structure of magnolia are formed from multiple pistils within a single flower, each one developing into a small fleshy unit. The red seeds that dangle from mature magnolia fruits on silk-like threads are the individual seeds emerging from those units.
Understanding this progression, from flower structure through pollination to fruit type, turns the litter under any tree from a nuisance into a readable record of its reproductive biology.
Why Flower Anatomy Matters in Practice
Flower anatomy might sound like textbook botany with no practical application. The opposite is true. Understanding tree reproductive biology informs decisions about cultivar selection, site management, client education, and pest timing.
Cultivar Selection and the Sex Problem
For dioecious species, specifying sex at planting is a management decision. Male ginkgo, male ash, and male poplar cultivars are standard specifications in street tree planting to avoid fruit litter. The unselected ginkgo seedling at the top of this guide had roughly a 50/50 chance of being female; a named male cultivar would have removed that risk before it was planted. But several decades of preferring male clones in urban forestry has created a documented side effect: elevated pollen loads in cities where the tree canopy is disproportionately male. Tom Ogren developed the Ogren Plant Allergy Scale (OPALS), rating plants 1 to 10 for allergenic potential, and argued that urban allergy seasons have been intensified by this “botanical sexism.” The USDA adopted OPALS for its plant database.
The science is more nuanced than the headline. Some researchers question whether the proportion of male-to-female urban trees is actually as skewed as Ogren claims, and others note that major allergens like alder and birch are monoecious (every tree produces pollen regardless of cultivar selection). But the underlying principle is sound: planting only male trees of dioecious species eliminates fruit litter while maximizing pollen output. A balanced approach mixes sexes and favors insect-pollinated species in areas where allergy is a concern, because insect-pollinated trees produce far less airborne pollen.
Fruit Litter as a Management Issue
Sweetgum balls, crabapple fruit, mulberry staining, and ginkgo sarcotesta are all consequences of female reproductive success. When clients ask for a tree that “doesn’t make a mess,” they are asking about flower biology whether they know it or not. Growth regulators containing ethephon or NAA (naphthaleneacetic acid) can suppress fruit set on some ornamental trees, but these are temporary, annual treatments, not permanent solutions. Understanding which trees are dioecious (where sex selection at planting solves the problem permanently) versus monoecious (where every tree will fruit and the only options are sterile cultivars or chemical suppression) helps you advise clients before the tree goes in the ground.
Pollination and Fruit Production
Arborists advising on fruit tree establishment need to understand cross-pollination requirements. A client who plants a single Honeycrisp apple and expects bushels of fruit will be disappointed. The conversation about pollinizer placement (which compatible cultivar, how close, will a crabapple work) starts with self-incompatibility. It applies to apples, pears, sweet cherries, Japanese plums, and some blueberries. European plums and sour cherries are largely self-fertile, a distinction worth knowing when advising on small-lot orchards where space for a second tree is limited.
Bloom Timing as a Management Cue
Flowering is the most conspicuous phenological event in the landscape, and it serves as a management trigger throughout the growing season. Cherry bloom in the Puget Sound lowlands signals the opening of the ornamental pest season. Crabapple bloom marks the approximate timing for codling moth egg-laying. Lilac bloom has been used for decades as an indicator for multiple pest emergence windows. HFG tracks bloom timing in growing degree days (GDD base 32) precisely because flower phenology is a reliable, observable cue that anchors the rest of the seasonal management calendar.
Flowers as Disease Entry Points
Some of the most damaging tree diseases enter through flowers. Fire blight (Erwinia amylovora) infects apple, pear, and ornamental rosaceous trees primarily through open blossoms, where the bacteria colonize the stigma and style before moving into the branch. Bloom-period sprays (streptomycin or copper) are timed to protect open flowers. Brown rot in stone fruit follows a similar infection pathway. Understanding that flowers are not just aesthetic features but active infection courts changes how you think about spray timing and sanitation. Dead-heading infected blossoms is not cosmetic care; it is disease management.
What to Notice This Season
Step outside during the next bloom cycle and look at trees differently. In February and March, find an alder or birch and examine the catkins: those are staminate (male) flowers, imperfect and wind-pollinated, the source of the yellow dust on your car. Look for the small, woody pistillate (female) structures on the same branch. That is a monoecious tree.
In April, look at a cherry or apple in bloom. Pull a single flower apart and find the four whorls: the green sepals at the base, the white or pink petals, the ring of stamens with yellow anthers, and the pistil at the center. That is a complete, perfect flower.
On a Douglas-fir, find the small reddish male cones near branch tips and the familiar woody female cones with their three-pointed bracts. That is a gymnosperm reproducing without flowers.
Every tree you pass is running a reproductive strategy shaped by 300 million years of evolution. Now you can read it.
Sources
- Reproductive Plant Parts. Oregon State University Extension. Clear overview of flower structure, complete/incomplete, perfect/imperfect classification, pollination types.
- Flower Parts (HO-173). Purdue University Extension. Concise illustrated reference on flower anatomy terminology.
- Parts of a Flower: An Illustrated Guide. American Museum of Natural History. Visual explainer of flower morphology.
- Angiosperm Flowers. Biology LibreTexts. Detailed botanical anatomy: whorls, ovary position, monocot/dicot patterns.
- Life Cycle of a Conifer. Biology LibreTexts. Gymnosperm cone anatomy, pollination drop mechanism, multi-year seed development.
- Botanical Sexism Cultivates Home-Grown Allergies. Scientific American. Tom Ogren’s argument on male-clone urban planting and allergy burden.
- Have Cities Really Made Allergies Worse by Planting Mostly Male Trees?. Slate. Counter-analysis of the botanical sexism thesis.
- Virginia Tech Dendrology Factsheets. Virginia Tech. Species-level flower and reproductive descriptions for over 1,000 North American trees.
- Pollination Mechanisms and Plant-Pollinator Relationships (M402). University of Missouri Extension. Pollination biology, wind vs. insect strategies, pollinator ecology.
- ISA BCMA Chapter 2: Tree Biology. International Society of Arboriculture. Key terms: angiosperm, gymnosperm, ethylene, abscission zone, epidermis.
- USDA Plants Database — Acer macrophyllum. USDA Natural Resources Conservation Service. Taxonomy, distribution, and reproductive classification (polygamous) for bigleaf maple.
Pesticide and growth regulator mentions are for educational context. Always read and follow the product label. Label directions supersede all other recommendations.
Get seasonal bloom timing, GDD tracking, and pest emergence alerts for the Puget Sound lowlands in the Field Brief, delivered free to your inbox each month.